
嗅刺激诱导的c-fos原癌基因在大鼠脑内的表达
摘 要 目的 为了解嗅刺激引起中枢神经系统兴奋的区域及程度。方法 以醋酸戊酯多次刺激大鼠,1h后灌注固定处死动物。以c-fos探针的ABC免疫细胞化学法,观察分析c-Fos蛋白在大鼠脑内标记区。结果 中枢系统内c-Fos蛋白标记区十分广泛,尤以脑桥以上脑区为多。除嗅觉通路诸结构外,边缘系统内也有广泛标记,提示存在嗅情绪反应、嗅内脏与嗅躯体反应及嗅记忆等多神经回路的活动。其次,与注意及中枢功能状态调节有关的脑区也有广泛标记。上丘深层、下丘及脑桥核等为强标记,推测与感觉和运动的整合活动有关。结论 用嗅刺激诱导c-fos原癌基因表达,确可证实一种特异性嗅刺激可引起大鼠中枢神经系统内泛脑网络上的广泛运动。
大鼠属嗅敏动物,其嗅脑和边缘系统相当发达。在选择配偶、御敌、觅食及其他探究性行为当中,大鼠嗅觉发挥着十分重要的作用。一种感觉刺激进入中枢神经系统(CNS)后,决非仅激活相应的感觉系统,而是涉及十分广泛的脑区和神经回路,即引起泛脑网络的多重活动[1]。以嗅球注射HRP显示嗅球传入及传出联系,不能全面反映CNS内整个神经活动的状况[2]。而以c-fos原癌基因作为神经活动与形态定位相结合的表达水平探针,其空间分辨力可达细胞水平[3~5]。本研究在大鼠体内以嗅刺激诱导c-fos原癌基因表达,试图以免疫细胞化学法观察与嗅刺激相关的脑区,分析其涉及的各种神经回路,全面认识嗅刺激引起的多重神经活动。
1 材料和方法
16只SD大鼠,雄性,体重150 g左右。其中嗅刺激实验组8只,嗅刺激物为浸透醋酸戊酯(香蕉香精)的棉球;对照组8只给予蒸馏水棉球,其中4只动物免除一抗步骤。
为避免无关刺激的干扰,预先彻底清洗鼠笼,将鼠放置在安静而无强光刺激的环境中,1 h后开始嗅刺激。嗅刺激时,先轻轻将刺激物放置在鼠笼顶刺激60 s,然后移除嗅刺激物2 min;照此反复嗅刺激10次。
最后一次嗅刺激后60 min(c-fos表达高峰期)[6],经升主动脉输入灌注固定液处死动物。在2%戊巴比妥钠(60 mg/kg)腹腔麻醉下,经升主动脉输入冲洗液(0.05 mol/L生理盐水磷酸缓冲液)100~150 ml,接着输入灌注固定液(4%多聚甲醛的0.05 mol/L磷酸缓冲液,pH 7.4)800ml,随即取脑和颈髓,置于固定液内过夜。经蔗糖磷酸缓冲液防冰晶处理,行冰冻切片,厚度为50μm。兼取冠状切片与矢状切片。
免疫细胞化学实验按切片漂浮法ABC程序进行。一抗为抗c-fos的绵羊多克隆抗体(美国CRB公司)。经预试验最佳一抗稀释度为1∶6 000。二抗与三抗使用Vectastain Elite ABC药盒。在一抗孵育前,以正常兔血清封闭非特异性蛋白结合位点1 h。在三抗孵育前,以3‰ H2O2的甲醇液淬灭内源性过氧化酶活性20 min。一抗孵育时间为70 h(4℃)+2 h(室温),二抗和ABC三抗孵育时间均在2 h以上。二抗与三抗稀释依药盒说明进行。在DAB呈色后贴片,以0.25% 的硫堇复染,常规脱水、透明及封片。c-Fos蛋白标记表现为深棕色细胞核。依大鼠脑图谱[7,8]识别标识的神经结构。依标记神经元胞核数目和呈色强度,分为极强(++++)、强(+++)、次强(++)及弱(+)四级。
2 结果
嗅刺激引起动物的一般行为反应 给予生理盐水棉球刺激,未引起动物注意及行为反应。给予醋酸戊酯棉球刺激,动物反应虽有个体差异,但均有明显的行为改变。有的动物从困倦中惊醒,以前爪洗面,打喷嚏,竖起躯体探究刺激物,然后远离刺激物;有的动物在第三次嗅刺激时才引起注意,但无明显探究行为。 在后几次嗅刺激时,动物正常饮水或进食,同未给刺激物的间歇期行为。
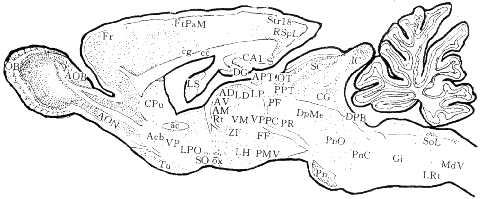
嗅刺激诱导c-fos在大鼠脑内的表达 c-Fos蛋白在CNS内的表达区域十分广泛。在低倍镜下观察矢状切片,嗅觉传导通路诸灰质结构均有强标记;大脑皮质额叶前部标记细胞浓密,而背侧海马仅稀疏标记;下丘脑标记区十分广泛但多不浓密,背侧丘脑总体标记密度明显不如下丘脑,大部见于其背侧边缘区;上丘深层、下丘及脑桥核的标记浓密,而脑桥以下标记结构明显减少。在冠状切片上,大脑半球内侧皮质、嗅周皮质以及杏仁核区的标记十分浓密(图1)。
fig 1 Diagram of expressed structures of c-Fos protein induced by an olfactory stimulus
OB: olfactory bulb; AOB: accessory olfactory bulb; AON: anterior olfactory nu; Fr: frontal cortex; FrPaM: frontoparietal cortex,motor area; cg: cingulum; CPu: caudate putamen; ac:anterior commissure; Acb: accumbens nucleus; VP:ventral pallidum; Tu:olfactory tubercle; cc:corpus callosum; LS:lateral septal nu; LPO: lateral preoptic area; AD:anterodorsal thalamic nu; AV:anteroventral thalamic nu; AM:anteromedial thalamic nu; Rt:reticular thalamic nu; SO:supra-optic hypothalamic nu; ox:optic chiasm; DG:dentate gyrus; LD: laterodorsal thalamic nu; ZI:zona incerta; CA1:field CA1 of Ammon's horn; APT: anterior pretectal area; LP:lateroposterior thalamic nu: VM:ventromedial thalamic nu: LH:lateral hypothalamic area: Str18: striate cortex,area 18; RSpL:retrosplenial cortex; S: subiculum; PF:parafascicular thalamic nu: VPPC:ventroposterior thalamic nu,parvocellular part; FF:field of Forel; PMV:premammillary nu; OT:nu of the optic tract; PPT:posterior pretectal area; PR:prerubral area; DpMe: deep mesencephalic nu; IC: inferior colliculus; SC: superior colliculus; CG: central grey; DPB:dorsal parabrachial nu; PnO:pontine reticular nu,oral part; PnC: pontine reticular nu,caudal part; Pn: pontine nu; Gi:gigantocelluar nu; Sol: nu of the solitary tract; LRt: lateral reticular nu: MdV: reticular nu medulla,vent
分脑区观察(结果):(1) 嗅觉传导通路上各灰质结构的标记(图2A~D):除嗅内区标记密度(++)明显下降外,其余灰质结构均有强标记(++++)。在嗅球中,以粒细胞层和僧帽细胞层标记最浓密,刷状细胞和小球周细胞次之。(2)大脑皮质内的标记(图2E~G):标记浓密的皮质区包括眶额皮质、扣带皮质前部、嗅周皮质、颞皮质及岛皮质等(均为++++)。海马结构总体标记稀疏,背侧海马(+)更不如腹侧海马(++),标记细胞仅见于锥体细胞层。(3) 端脑内诸神经核的标记(图2H,I):最浓密为屏状核和杏仁核复合体(+++~++++)。隔核标记强度(+++)明显大于海马结构。在纹状体中,尾壳核标记(++)主要位于其背外侧部,苍白球内未见明确标记。腹侧苍白球(+++)、纹状基(++)以及无名质(++)等腹侧结构标记强度稍大于尾壳核背部(++)。在诸前脑基底结构内标记广泛但不很浓密。(4) 间脑内各神经核的标记(图2J,K):在矢状切片上观察,背侧丘脑内最显著标记区靠近其背侧缘和后缘区,以丘脑室旁核(++~+++))和束旁核(+++)较为突出;在丘脑髓纹后方的丘脑网状核(+++)和未定带(+++)标记也较显著。在冠状切片上观察,丘脑中线核群、板内核群、前核群、内侧背核外侧部均有稀疏标记(+)。位于内侧背核腹侧的丘脑腹内侧核(+++),标记区虽小但密度相对较大。缰核标记(++~++++)十分明显。腹外侧膝状体小细胞部(++)和内侧膝状体的下外侧边缘区(+)也有少量标记。值得注意的是,丘脑外侧核群,尤其是腹侧组未见标记。在下丘脑标记区十分广泛,以下丘脑前核(+++)、腹内侧核(+++)、穹隆周核(+++)、乳头体核(++)及下丘脑外侧区(++)较为显著。相反,属于神经分泌性核团,如下丘脑室旁核(++)、视上核(+)、弓状核(+)、结节外侧核(++)等标记并不突出。(5) 脑干与脊髓内的标记(图2L~O):在矢状切片上观察,十分醒目的标记区有背侧的顶盖前区核团、上丘深层、下丘、腹侧的脑桥核(均为++++)。属于脑干网状结构内侧区的各个网状核,在脑桥以上具有连续标记。在冠状切片上观察,所谓边缘中脑区,如导水管周围灰质、脚间核、被盖腹侧区、被盖背侧核等均有明确标记(均为++)。脑干上部的中缝核群(+)、脑桥背外侧的臂旁核(++)和蓝斑(+++)等也有明确标记。此外,三叉神经感觉核群(+)、孤束核外侧部(++)、外侧网状核(++)、最后区(+),以及颈髓后角边缘核(+++)和胶状质(++)等,也有强度不等的标记。
3 讨论
本实验结果表明,用嗅刺激诱导c-fos原癌基因表达,确能观察到一种新异的嗅刺激可引起嗅敏动物脑内广泛而又兴奋程度不一的变化。以往对大鼠嗅球神经联系的辣根过氧化物酶(HRP)研究[2]所发现的CNS内各种神经回路的绝大部分标记结构[9~15],均可通过c-fos原癌基因表达得到证实。
对于嗅敏动物大鼠,嗅刺激可引起其脑边缘系统的广泛神经活动,这可能与嗅情绪反应及行为反应有关。属边缘系统的脑区,在端脑,包括眶额皮质、扣带皮质、嗅内区、海马结构、杏仁核复合体及隔核等;在间脑主要是下丘脑;在中脑为边缘中脑区诸结构,即所谓“边缘系统-中脑回路”的尾侧极,属于脑干网状结构的旁正中亚区,其又与孤束核及臂旁核等内脏感觉中枢核联系在一起。本研究表明,c-Fos蛋白标记区囊括了上述诸神经结构。
根据边缘系统诸结构的功能与c-Fos蛋白标记强度分析,与嗅情绪反应有关的眶额皮质、扣带皮质前部及杏仁复合体均具有浓密标记,提示醋酸戊酯可引起大鼠很强的情绪反应。而主要参与嗅躯体反应和嗅内脏反应的脑区,如下丘脑诸核团、丘脑内侧背核、缰核、脚间核、孤束核及最后区等则标记强度较弱,推测醋酸戊酯属于与大鼠觅食及性行为无关的嗅刺激,故不能引起其强烈的行为反应。包括嗅内区、海马结构、乳头体核、丘脑前核等近记忆回路诸结构的标记强度大都不很显著,这可能与醋酸戊酯对大鼠属无关刺激有关,10次重复刺激并未引起动物深刻记忆。
本实验表明,特异性嗅刺激可引起脑内非特异性功能状态系统广泛调变。脑干上部中缝核群(5-HT能神经元)与蓝斑(NE能神经元)属于中枢功能状态调节系统。脑干内侧网状核群、丘脑非特异性核群(中线核群、板内核群、丘脑网状核)、未定带、前脑基底结构及广泛的大脑皮质区,均属上行网状激动系统。而上述脑区均有c-Fos蛋白标记,但标记强度均不明显。应当指出,嗅刺激引起鼠脑内如此广泛的非特异性功能状态调节系统调变的上述绝大部分结构均在脑桥平面以上,其意义有待进一步研究。
嗅觉与味觉的密切相关性得到进一步证实,副嗅球、杏仁皮质后内侧核、杏仁中央核、终纹床核、无名质、丘脑腹内侧核(味觉区)、臂旁核(味觉区)及孤束核外侧部等,都有明确的c-Fos蛋白标记。这些脑区的激活源自醋酸戊酯对鼻腔犁鼻器的刺激。
对其他c-Fos标记区的可能解释:(1)屏状核靠近嗅周皮质,实验组动物均有浓密标记,这至少说明此核与嗅觉有密切关系。(2) 某些脑区的标记可能与嗅刺激引起探究刺激源的活动有关。例如,顶盖前区在控制眼跟踪运动中起作用。上丘深层是重要的皮质下感觉运动整合区,使动物在接受多种刺激后,眼、头及躯干联合协调运动而转向刺激源。下丘也是多种感觉信息与运动信息的整合部位。属于小脑前核的脑桥核和外侧网状核至少也涉及感觉与运动信息的整合功能。除外侧网状核外,上述诸结构的浓密标记均提示嗅刺激引起探究行为的感觉与运动整合活动占有很大的比重。而真正执行躯体和眼球运动的神经结构,却没有任何标记。(3) 醋酸戊酯除兴奋鼻腔嗅粘膜外,还能刺激鼻腔呼吸道粘膜,引起三叉神经感觉核群的兴奋。而三叉神经脊束核又与颈髓后角边缘核及胶状质是同源相连续结构,因此后者也有标记。(4) 下丘脑有关神经分泌核团没有明显标记,推测与醋酸戊酯属对大鼠为非生理性无关嗅刺激有关;也可能与本实验仅选择c-fos表达60 min,表达时间不够长有关。
杨天祝为通讯作者
作者单位:杨天祝 葛莉 (河北医科大学 河北省基础医学研究所神经生物学研究室,石家庄 050017)
万选才 (中国医学科学院中国协和医科大学基础医学研究所,北京,100730)
参考文献
1 万选才.脑的理论模式.生理科学进展,1987,18(2):181~184
2 刘东刚,杨天祝,杨 华.大鼠嗅球的纤维联系.河北医学院学报,1988,9(1):1~4
3 Dragunow M,Faull R.The use of c-fos as a metablic marker in neuronal pathway tracing.J Neurosci Methods,1989,29:261~265
4 Bullitt E.Expression of c-fos-like protein as a marker for neuronal activity following noxious stimulation in the rat.J Comp Neurol,1990,296:517~530
5 Anton F,Herdegen T,Peppel P,et al.c-Fos-like immunoreactivity in rat brainstem neurons following noxious chemical stimulation of the nasal mucosa.Neuroscience,1991,41(2/3):629~641
6 Wan XST,Liang F,Moret V,et al.Mapping of the motor pathways in rats: c-fos induction by intracortical microstimulation of the motor cortex correlated with efferent connectivity of the site of cortical stimulation.Neuroscience,1992,49(2):749~761
7 Paxinos G,Watson C.The rat brain in stereotaxic coordinates.Sydney: Academic Press,1986.15~54
8 王平宇,张风真.大鼠脑读片提要及图谱.西安:西安大学出版社,1995.23~219
9 河北新医大编写组.人体解剖学.北京:人民卫生出版社,1978.1262~1268
10 张培林.神经解剖学.北京:人民卫生出版社,1987.477~491
11 Williams PL,Warwick R.Functional neuroanatomy of man.Philadelphia: W.B.Saunders Company,1975.936~943
12 Nieuwenhuys R,Voogd J,Huijzen Chr van.The human central nervous system.Berlin: Springer-Verlag,1978.181~211
13 Brodal A.Neurological antomy.3rd ed.New York: Oxford University Press,1981.640~697
14 Paxinos G.The human nervous system.San Diego: Academic Press,1990.978~998
15 Kandel ER,Schwartz JH.Principles of neural science.3rd ed.New York: Elsevier,1991.516~518




- 两性
- 男人
- 女性
- 母婴
|
・ 处女座的特点 ・ 处女座最佳配对星座 ・ 2010年处女座运势 ・ 处女座女人的爱情 ・ 如何追处女座女人 ・ 处女座女人的特点 ・ 处女座女人 ・ 处女座男人喜欢的女人 ・ 如何对付处女座男人 |

|
・ 怎样看待遗精 ・ 什么是滑精 ・ 什么是梦遗 ・ 什么是干燥性闭塞性龟头炎? ・ 前列腺炎检查 ・ 包皮手术过后多久可以性生活 ・ 早泄是不是跟包皮过长有关? ・ 早泄等于射精过快吗? ・ 体外射精有什么害处 |
|
・ 女性经期切记将绿茶挡在门外 ・ 生命中的一次婚外恋 ・ 一个流氓和妓女的故事 ・ 最唯美的10首中国情诗 ・ 娇妻玩合租 结果引火烧身 ・ 男人必须了解女人的一些事 ・ 当女朋友被领导叫去陪酒 ・ 易让男人退避三舍的10类女人 ・ 老男人为什么招小女人的喜欢? |
|
・ 春季合理喂养婴儿健康指南 ・ 如何正确使用空调保证健康 ・ 让宝宝接受保姆的三大招 ・ 哪些产妇需做会阴侧切 ・ 导致分娩时难产4因素 ・ 看美国准妈人性化孕产经历 ・ 准妈妈如何预防春季感冒? ・ 胎盘和脐带的功能与重要性 ・ 烟、酒和咖啡对胎儿的影响 |